Растения и животные
ЭНЦИКЛОПЕДИЯ
Как размножаются растения
Как и все живые организмы, растения размножаются. Существует три способа размножения растений — вегетативный, бесполый и половой. При вегетативном способе новая особь образуется из части вегетативных органов растений, т. е. листа, стебля или корня. Иногда новая особь возникает даже из отдельной клетки того или иного вегетативного органа растения. При бесполом размножении у растений образуются особые клетки (споры), из которых вырастают новые самостоятельно живущие особи, сходные с материнской. Этот способ размножения свойствен некоторым водорослям (рис. 1) и грибам (см. ст. «Грибы»). Половое размножение принципиально отличается от вегетативного и бесполого. Половой процесс в растительном мире крайне разнообразен и часто очень сложен, но по существу сводится к слиянию двух половых клеток — гамет, мужской и женской.
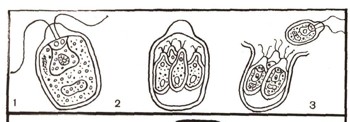
Рис. 1. Бесполое размножение хламидомонады: 1 — хламидомонада; 2—образование зооспор; 3 — выбегание зооспор.
Гаметы возникают в определенных клетках или органах растений. В некоторых случаях гаметы одинаковы по размерам и форме, обе имеют жгутики и потому подвижны. Это изогамия (рис. 3). Иногда они несколько отличаются друг от друга размерами. Это гетерогамия (рис. 2). Но чаще — при так называемой оогамии — размеры гамет резко различны: мужская гамета, называемая сперматозоидом, небольшая, подвижная, а женская — яйцеклетка — неподвижная и крупная (рис. 4). Процесс слияния гамет называется оплодотворением. Гаметы имеют в своем ядре по одному набору хромосом, а в образовавшейся после слияния гамет клетке, которая называется зиготой, число хромосом удваивается (см. ст. «Клетка»). Зигота прорастает и дает начало новой особи.
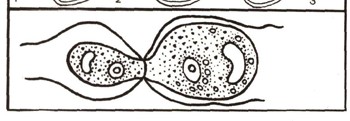
рис. 2. Гетерогамия у хламидомонады. Обе гаметы подвижны, но отличаются размерами.
Половой процесс осуществляется у растения в определенное время и на определенном этапе развития. На протяжении своего развития растение может размножаться также и бесполым путем (с образованием спор) и вегетативно.
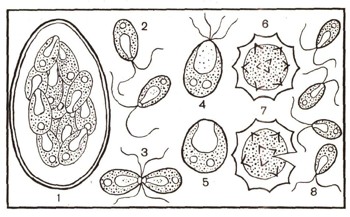
Рис. 3. Изогамия у хламидомонады: 1 — образование гамет; 2 — гаметы; 3 — слияние гамет; 4 — зигота (видны жгутики); 5 — зигота со сброшенными жгутиками; 6,7,8 — прорастание зиготы и образование четырех новых особей хламидомонады.
Половое размножение возникло в растительном мире в процессе эволюции. У синезеленых водорослей его еще нет. Они размножаются только вегетативно, путем деления клетки на две. У большинства водорослей и грибов, а также у всех высших наземных растений половой процесс отчетливо выражен. Половое размножение очень важно для организма, так как благодаря слиянию отцовской и материнской клеток создается новый организм, который имеет большую изменчивость, лучше приспособлен к условиям окружающей среды.
Наиболее прост процесс полового размножения у одноклеточных водорослей, например у хламидомонад. Хламидомонада размножается как бесполым, так и половым путем. При бесполом размножении хламидомонада теряет жгутики и делится на 2, 4 (реже 8) клетки-споры. Каждая из них снабжена двумя жгутиками. Это зооспоры. После разрушения оболочки клетки, внутри которой они сформировались, зооспоры разбегаются и дорастают до размеров материнской клетки (рис. 1).
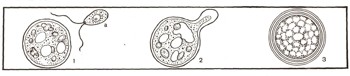
Рис. 4 Оогамия у хламидомонады: 1 — неподвижная яйцеклетка, а — сперматозоид; 2 — оплодотворение (слияние сперматозоида с яйцеклеткой); 3 — зигота, покрывшаяся толстой оболочкой.
При половом размножении (рис. 3) содержимое клетки хламидомонады делится и образуется большое число гамет (32 или даже 64). Потом оболочка материнской клетки прорывается, и гаметы, имеющие по два жгутика, выходят в воду, плавают, соединяются попарно своими носиками, где расположены жгутики, и, наконец, полностью сливаются друг с другом. У большинства хламидомонад трудно отличить, какие гаметы мужские, какие женские. Они одинаковы по форме и одинаково подвижны. Однако есть некоторые виды хламидомонад, которые образуют неподвижные крупные женские гаметы (яйцеклетки), а другие особи — мелкие подвижные мужские гаметы (сперматозоиды). После слияния гамет жгутики исчезают, образуется зигота, которая сразу же покрывается оболочкой (рис. 4).
Через некоторое время зигота прорастает. Первое деление ее ядра редукционное — особое деление ядра, при котором число хромосом в клетке уменьшается вдвое (см. ст. «Клетка»). В результате второго деления каждого из ядер образуются 4 клетки с одним набором хромосом в их ядрах. Оболочка зиготы лопается, и новые клетки выходят в воду, плавают при помощи двух жгутиков. Достигнув размеров материнской клетки, они могут снова размножаться бесполым и половым путем.
Период от появления гаметы и до образования новых гамет называют циклом развития растения. У некоторых многоклеточных водорослей обе половые клетки неподвижны. Так, у спирогиры при половом процессе содержимое одной клетки переливается в другую, где и происходит слияние их цитоплазмы, ядер и образуется зигота. У других многоклеточных водорослей процесс полового размножения более сложен.
Этот процесс очень разнообразен у наземных растений. У мхов, папоротников, голосеменных, например у хвойных, а также у цветковых растений он происходит по-разному. В связи с выходом из воды на сушу у мхов, папоротников, хвощей, плаунов и у семенных растений сильно усложнилось не только строение, но и процесс размножения. У них, как и у многих водорослей, наблюдается правильное чередование бесполого и полового поколений. Зигота прорастает без редукционного деления, и развивающаяся из нее особь имеет двойной набор хромосом.
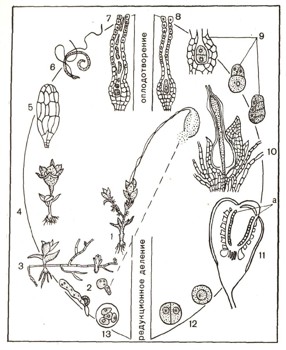
Рис. 5. Развитие мхов: 1 — взрослое растение; 2 — прорастание споры (2 фазы); 3 — предросток с молодыми растениями мха; 4 — мужское растение с антеридиями; 5 — антеридий; 6 — сперматозоид; 7 — архегоний, готовый к оплодотворению; 8 — оплодотворенный архегоний; 9 — первые стадии развития спорогона; 10 — молодой спорогон внедрился ножкой в верхушку стебля; 11 — зрелая коробочка с клетками (а), из которых в дальнейшем образуются споры; 12 — подготовка клеток к редукционному делению; 13 — гаплоидные споры.
Это бесполое поколение, так как на таком растении образуются споры. При формировании их происходит редукционное деление, в результате которого спора получает один набор хромосом. Прорастающая спора дает начало половому поколению — организму, который образует половые клетки — гаметы. Все клетки этой особи несут один набор хромосом. Образующаяся в результате оплодотворения гамет зигота снова прорастает и дает бесполое поколение (с двойным набором хромосом). В цикле развития растения может преобладать половое (мхи) или бесполое (остальные высшие растения) поколение.
Рассмотрим цикл развития мха кукушкин лен (рис. 5). Стебли этого мха небольшие, крепкие, с многочисленными мелкими, узкими, жесткими листьями. На верхушке некоторых из этих стеблей развиваются коробочки, сидящие на удлиненной ножке и покрытые колпачком, как капюшоном (1). Коробочка на ножке называется спорогоном. В самой коробочке, покрытой крышечкой, образуется масса спор. Они мелкие, как пыль. При их образовании происходит редукционное деление, и споры
получают по одному набору хромосом (гаплоидные споры). После их созревания колпачок сбрасывается, крышечка коробочки отскакивает, и споры высыпаются наружу. Споры попадают на почву и при влажной погоде прорастают (2). Образуется зеленая ветвистая многоклеточная нить, стелющаяся по влажной поверхности почвы, а погруженные в почву бесцветные нити всасывают почвенные растворы. Зеленая нить называется предростком (3). На предростке образуются почки. Из почек развиваются новые стебли кукушкина льна.

На верхушках одних стеблей появляются многоклеточные небольшие кувшинообразные выросты, сидящие на небольшой ножке. Это женские половые органы, или архегоний. В их нижней расширенной части помещается одна неподвижная яйцеклетка. На верхушках других стеблей мха вырастают многоклеточные, но одностенные удлиненные мешочки — антеридии (4, 5). Внутри них образуются многочисленные мелкие мужские гаметы — сперматозоиды (6). Во время дождя или обильной росы мешочки лопаются вверху, и из них в слизистой массе выступает множество сперматозоидов, снабженных двумя жгутиками, с помощью которых они двигаются к верхушке тех стеблей кукушкина льна, где находятся архегоний. Проникнув через шейку архегония внутрь, сперматозоид сливается с яйцеклеткой (7, 8). В результате образуется зигота, которая прорастает без редукции хромосом здесь же, на верхушке стебля кукушкина льна, образуя бесполое поколение — спорогон, состоящий из коробочки и ножки. Ножка спорогона внедряется в ткани стебля и высасывает из него питательные вещества (9, 10), В коробочке спорогона образуются споры (11, 12, 13). Таков цикл развития мхов. У них преобладает половое поколение (само растение мха).
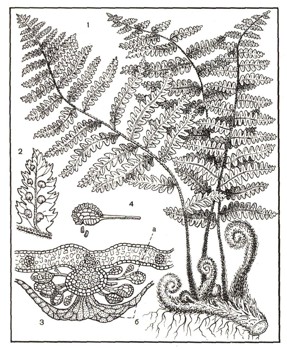
Рис. 7. Папоротник: 1 — внешний вид папоротника (бесполое поколение); 2 — долька листа с нижней стороны (видны сорусы, одетые покрывалом); 3 — разрез соруса, а — спорангии, б — покрывало; 4 — отдельный спорангий, из которого высыпаются споры.
Теперь рассмотрим цикл развития у папоротника щитовника, распространенного по тенистым местам в лиственных лесах (рис. 6, 7). Из верхушки его подземного корневища ежегодно вырастает пучок перистосложных листьев (1). На нижней поверхности листьев вдоль средней жилки легко заметить кучки спорангиев — так называемые сорусы, закрытые покрывалом, напоминающим в поперечном разрезе раскрытый зонтик (2, 3). Двояковыпуклый спорангий имеет вид чечевицы и расположен на ножке. Внутри спорангия масса мелких спор, возникших в результате редукционного деления.
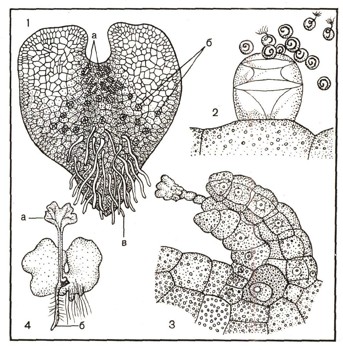
Рис. 8. Половое размножение папоротника: 1 — заросток, а — архегонии, б — антеридии, в — ризоиды; 2 — из зрелого антеридия выходят сперматозоиды; 3 — архегонии, готовый к оплодотворению; 4 — заросток с молодым спороцоном, а — первый лист, б — корешок.
В сухую погоду, когда споры уже созрели, спорангий вскрывается (4). Высыпавшиеся от резкого толчка споры рассеиваются и попадают на поверхность почвы. Попав в благоприятные условия — тепло и влагу, спора прорастает и образует очень маленькую (2—5 мм в диаметре) тонкую зеленую пластинку сердцевидной формы — заросток (рис. 8). Своей нижней поверхностью заросток плотно прижимается к земле благодаря ризоидам, всасывающим из почвы растворы минеральных солей. Заросток папоротника обоеполый: на его нижней поверхности расположены женские (архегонии) и мужские (антеридии) половые органы. Заросток и представляет собой половое поколение папоротника. Во время дождя или обильной росы многожгутиковые сперматозоиды выходят из антеридия в воду и направляются к архегонию. Там происходит оплодотворение, после чего получается зигота — клетка с двойным набором хромосом. Она прорастает здесь же, на заростке, и образуется зародыш. Разрастаясь все больше и больше, он образует все части взрослого растения: стебель, лист, корни. Затем на нижней поверхности листа взрослого растения снова появляются сорусы со спорангиями.
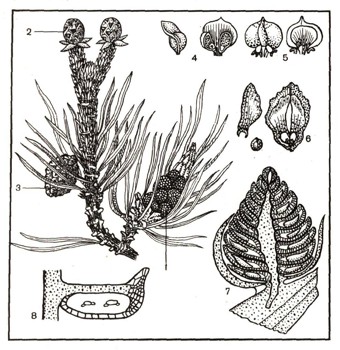
Рис. 9. Мужские и женские шишки сосны: 1 — собрание мужских шишек; 2 — молодая женская шишка на верхушке побега; 3 — прошлогодняя женская шишка; 4 — чешуйки мужской шишки (вид сбоку и снизу); 5 — чешуйки женской шишки (вид с наружной и внутренней стороны); 6 — чешуйка зрелой женской шишки с двумя крылатыми семенами, отдельно изображены крылышко и семя; 7 — продольный разрез мужской шишки, на оси расположены чешуйки со спорангиями; 8 — отдельный спорангий со спорами (пылинками).
Таким образом, в цикле развития папоротника преобладает бесполое поколение, формирующее спорангии со спорами (сам папоротник). Половое поколение (заросток) имеет незначительные размеры и существует недолго. Оба поколения существуют раздельно, самостоятельно. Подобным же образом размножаются хвощи и плауны, которые вместе с папоротниками объединяют в класс папоротникообразных.
По-другому идет размножение у семенных растений. У них рассеиваются не споры, а семена. Однако и у этих растений тоже образуются споры, а также двоякого рода клетки полового размножения: мужские и женские.
У голосеменных, например у сосны, ели, образуются мужские и женские шишки (рис. 9). Мужские шишки собраны в тесные группы у основания побегов, развивающихся в этом году. Женские шишки сидят поодиночке сначала на верхушке побега, а затем вследствие роста побега оказываются у его основания. Мужская шишка состоит из чешуек, тесно сидящих на ее оси. На нижней поверхности чешуек расположены два спорангия. Внутри спорангия путем редукционного деления развивается огромное количество спор (пылинок). Содержимое каждой пылинки состоит из густой плазмы и ядра. Пылинка одета оболочкой, образующей два пузырчатых сетчатых воздушных мешка (рис. 10). Это приспособление способствует разносу ветром пылинок, высыпавшихся из лопнувшего пыльника. Пылинка прорастает в мужской заросток. При этом ядро ее делится, и образуются две быстро разрушающиеся клетки и две сохраняющиеся дольше клетки — более крупная вегетативная и более мелкая антеридиальная. В таком двуклеточном состоянии пылинка переносится ветром и попадает на поверхность женской шишки, где и происходит процесс оплодотворения.
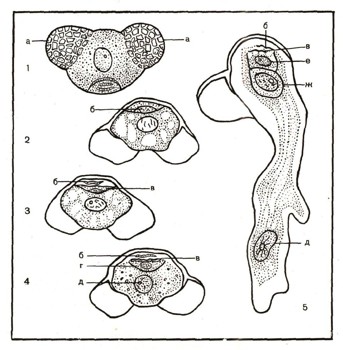
Рис. 10. Пылинка (спора) и развитие мужского заростка сосны: 1 — зрелая спора, а — воздушные мешки; 2—5 — прорастание споры и образование мужского заростка, б, в — рано исчезающие клетки заростка, г — антеридиальная клетка, д — вегетативное ядро (ядро клетки пыльцевой трубки), е — сестринская клетка, ж — генеративная клетка (при дальнейшем ее делении образуются два спермин — мужские гаметы).
Женская шишка состоит из мелких кроющих чешуек, в пазухах которых развиваются крупные мясистые семенные чешуи. У основания последних на их внутренней (верхней) стороне расположены по две овальные семяпочки (рис. 11). В верху семяпочки имеется маленькое отверстие — пыльцевход. В семяпочке одна из клеток, выделяющаяся крупными размерами, делится редукционно, в результате образуются четыре споры. Три из них отмирают, а четвертая приступает к делению. В результате многократного деления образующихся при этом клеток формируется женский заросток, занимающий середину семяпочки. На заростке образуются два небольших архегония очень упрощенного строения с маленькими шейками, в каждом из которых находится по одной яйцеклетке.
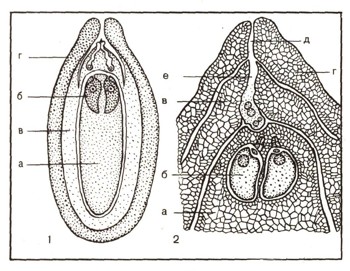
Рис. 11. Семяпочка сосны: 1 — продольный разрез семяпочки, о — женский заросток, б — архегонии, в — нуцеллус, г — покров; 2 — верхняя часть семяпочки в продольном разрезе при большем увеличении, а — женский заросток, б — яйцеклетка архегония, в — нуцеллус, г — покров, д — пыльцевход, е — пыльцевая трубка, проросшая через нуцеллус и достигшая женского заростка. В пыльцевой трубке видны 4 ядра (2 спермин, вегетативное ядро и ядро сестринской клетки).
Если теперь разрезать семяпочку вдоль, то можно увидеть, что заросток окружен содержимым семяпочки (нуцеллусом), который, в свою очередь, одет покровом семяпочки. Наверху осталось всего лишь маленькое отверстие — пыльцевход. Через него и попадает перенесенная ветром на верхушку семяпочки пылинка. Она втягивается внутрь семяпочки, где и прорастает на следующее лето. Пылинка образует пыльцевую трубку, внедряющуюся в нуцеллус и растущую по направлению к шейке одного из архегониев. В это же время антеридиальная клетка делится на две. Одна из образовавшихся клеток в дальнейшем разрушается, а другая (генеративная клетка) увеличивается в размере, делится и образует две половые клетки — мужские гаметы, или спермин, не имеющие жгутиков.
Следует, однако, отметить, что у более древних голосеменных растений (гинкго и саговников) имеются подвижные сперматозоиды. Это свидетельствует об их происхождении от папоротникообразных растений.
Достигнув архегония, пыльцевая трубка лопается, и один из спермиев попадает в архегонии и сливается с яйцеклеткой. Происходит оплодотворение, и образуется зигота. Другой спермий вскоре отмирает. Из зиготы формируется зародыш нового растения, питающийся за счет запасных веществ клеток женского заростка (эндосперма). Семяпочка становится теперь семенем. Семя одето плотной кожурой, в которую превратился покров семяпочки.
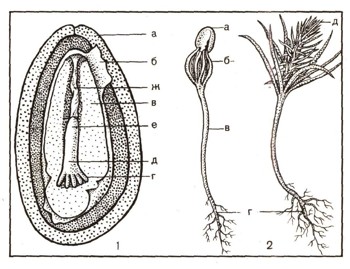
Рис. 12. Строение и прорастание семени сосны: 1 — строение семени, а — кожура (покров) семени, б — остаток нуцеллуса, в — эндосперм, в середине которого находится зародыш, г — семядоли зародыша, д — подсемядольное колено (зачаточный стебель) е — корешок, ж — подвесок; 2 — прорастание семени сосны, а — кожура семени, б — семядоли, в — подсемядольное колено, г — корень, д — побег с первыми листьями.
Семена созревают к осени. Они сидят в основании чешуи шишки. К осени на второй год своего существования шишка разрастается. Из зеленой она становится бурой, чешуи подсыхают, расходятся, семена выпадают и рассеиваются. Попав в благоприятные условия, семена прорастают и развиваются в новые растения (рис. 12).
В цикле развития сосны, так же как и у папоротникообразных, преобладает бесполое поколение. Половое поколение здесь еще более просто устроено. При этом оно потеряло способность к самостоятельной жизни и развивается внутри тканей бесполого поколения (женский заросток — внутри семяпочки, а мужской заросток — внутри пылинки).
Особенность размножения покрытосеменных (или цветковых) растений — образование цветка как специализированного органа, приспособленного к половому размножению (рис. 13). Внешняя часть цветка состоит из околоцветника, обычно в виде лепестков и чашелистиков. Но главную часть цветка составляют находящиеся в его центре пестик (или пестики) и тычинки, расположенные вокруг пестика. Тычинки состоят из тычиночных нитей и пыльников, а пестик — из одного или нескольких сросшихся между собой завернутыми внутрь краями плодолистиков. В образующейся при этом срастании полости скрыты расположенные обычно по краям плодолистиков одна или несколько семяпочек.
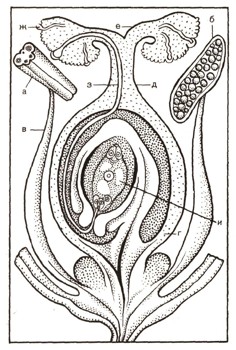
Рис. 13. Размножение у покрытосеменных (цветковых) растений. Продольный разрез цветка (чашелистики и лепестки удалены): а — тычинка, на ее поперечном разрезе видны гнезда пыльника; б — пыльник в продольном разрезе, видна пыльца; в — нить тычинки; г — завязь; д — столбик; е — рыльце пестика; ж — прорастающая на рыльце пылинка; з — пыльцевая трубка, проросшая через ткани рыльца, столбика и достигшая зародышевого мешка семяпочки; и — зародышевый мешок.
В нижней части пестик расширен. Это завязь. Кверху пестик утончается и образует столбик, который заканчивается по-разному устроенным рыльцем, служащим для улавливания и восприятия пыльцы. Плодолистики затем изменяются и принимают большое участие в образовании плода.
Как и у голосеменных, здесь центральную часть семяпочки занимает однородная ткань из живых клеток — нуцеллус. С внешней стороны нуцеллус прикрыт двумя, реже одним покровом. Наверху покровы не смыкаются. Здесь имеется отверстие — пыльцевход. Вскоре после образования нуцеллуса одна из его верхних клеток путем редукционного деления образует четыре споры. Одна из них сильно разрастается и приступает к делению, в результате образуется женский заросток — зародышевый мешок. Остальные три споры отмирают.
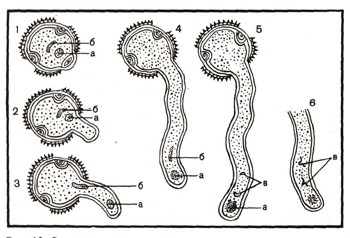
Рис. 14. Пыльца покрытосеменного (цветкового) растения и ее прорастание: 1 — пылинка, внутри видны округлое вегетативное ядро — ядро клетки пыльцевой трубки (а) и изогнутая генеративная клетка (б); 2 — через пору в наружной оболочке пылинки вытягивается пыльцевая трубка; 3 — вегетативное ядро опустилось в пыльцевую трубку; 4, 5 — генеративная клетка поделилась, образовались два спермия (в); 6 — зрелые спермин (в).
Женский заросток у покрытосеменных еще более упрощен по сравнению с голосеменными и состоит всего из 8 клеток. Образуется он так. Ядро споры делится на два. Расходясь к полюсам зародышевого мешка, они снова двукратно делятся. Теперь на полюсах уже по четыре ядра. Вскоре от каждой из этих четверок по направлению к центру мешка отделяется по одному ядру. Это полярные ядра. Здесь они сближаются, затем, сливаясь, образуют вторичное (центральное) ядро зародышевого мешка.
Ядра, оставшиеся на полюсах, облекаются цитоплазмой. Образуется по три клетки на каждом из полюсов. Противоположные от семявхода клетки называются антиподами. Три клетки, расположенные близ верхнего конца зародышевого мешка, не одинаковы. Средняя из них представляет собой яйцеклетку, а расположенные по бокам возле нее две меньшие клетки называются вспомогательными. Середина зародышевого мешка заполнена цитоплазмой и вакуолями с вторичным ядром в центре.
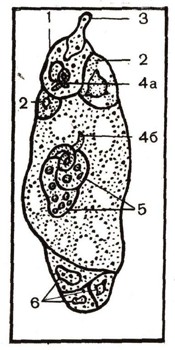
Рис. 15. Зародышевый мешок (женский заросток) покрытосеменного (цветкового) растения и двойное оплодотворение: 1 — яйцеклетка; 2 — вспомогательные клетки; 3 — вскрывшаяся пыльцевая трубка. Один из спермиев (4а) сливается с яйцеклеткой; 5 — полярные ядра, сливающиеся со вторым спермием (4б); 6 — три клетки в нижней части зародышевого мешка (антиподы).
В пыльнике тычинки, в каждом из четырех его гнезд образуются споры (пылинки). Они происходят из особых материнских клеток пыльцы в результате редукционного деления их. Содержимое пылинки состоит из крупного ядра и цитоплазмы (рис. 14). Пылинка окружена двумя оболочками: внутренней и внешней. Во внешней оболочке имеются отверстия или утонченные места. Еще в гнезде пыльника в каждой пылинке начинается формирование мужского заростка. Он еще более упрощен в сравнении с голосеменными. Ядро пылинки делится, и образуются две клетки: более крупная — вегетативная и более мелкая — генеративная. После этого пыльник вскрывается, пыльца из него высыпается и с помощью ветра, насекомых или воды, а у некоторых тропических растений при помощи птиц попадает на рыльце пестика. Этот процесс называется опылением.
Около 10% цветковых растений опыляется ветром. Цветки ветроопыляемых растений невзрачны. Они имеют околоцветник в виде пленок, чешуек; нередко он совсем отсутствует, например у злаков, осок, дуба, березы, осины, ольхи. Пыльца этих растений очень мелкая, с гладкой наружной оболочкой. Пыльцы образуется очень много, ведь ветер — ненадежный опылитель. Лишь небольшая часть пылинок попадает на рыльце пестика.
Большинство цветковых растений опыляется насекомыми: пчелами, осами, шмелями, бабочками, мухами. Насекомые посещают цветки из-за сладкого сока (нектара), который выделяется особыми нектарными желёзками, расположенными на лепестках, тычинках или на цветоложе. Венчики цветков насекомоопыляемых растений ярко окрашены и хорошо заметны издали. Пыльца у них более крупная, наружная оболочка пылинок имеет выросты в виде шипов, бугорков, и потому пыльца легко задерживается на рыльце пестика.
Рис. 16. Строение и прорастание семени у клещевины: 1 — проросток (2 стадии развития), а — оболочка семени, б — эндосперм, в середине которого находится зародыш, в — семядоли, г — подсемядольное колено — зачаточный стебель, д — корень; 2 — разрез семени.
Очень важно, чтобы пыльца не попадала на рыльце того же цветка. В случае самоопыления, как заметил еще Ч. Дарвин, получается более слабое потомство. У растений имеются различные приспособления, обеспечивающие перекрестное опыление, при котором пыльца попадает на рыльце другого цветка. Так, у ветроопыляемых растений цветки большей частью раздельнополые: одни цветки содержат только тычинки (тычиночные цветки), другие — только пестики (пестичные цветки). У насекомоопыляемых растений цветки, как правило, обоеполые, имеющие тычинки и пестики. Очень часто тычинки созревают и начинают высыпать пыльцу значительно раньше, чем полностью сформируется пестик. У многих растений пестики созревают раньше тычинок. У некоторых растений, например у примулы, медуницы, незабудки, тычинки и пестики не одинаковы по длине. Наиболее здоровое и сильное потомство вырастает из семян, образовавшихся в результате перенесения пыльцы из цветков с длинными тычинками на рыльце пестиков с длинными столбиками.
Попавшая на рыльце пестика пыльца прорастает (рис. 14). Вегетативная клетка, находящаяся внутри пылинки, разрастается и вытягивается в пыльцевую трубку, которая выходит через отверстие во внешней оболочке пылинки и продвигается в виде тонкой нити сквозь рыхлую ткань рыльца и стенок пестика к семяпочке. Через пыльцевход она направляется к зародышевому мешку.
Во время роста пыльцевой трубки в нее проникает генеративная клетка. Здесь она делится и образует две мужские гаметы (спермин). Достигнув зародышевого мешка, пыльцевая трубка, в которой находятся вегетативное ядро и два спермин, лопается, и содержимое ее изливается в зародышевый мешок (рис. 15). Один из спермиев сливается с яйцеклеткой. Образуется зигота. Второй спермий направляется в середину зародышевого мешка и сливается там со вторичным ядром. Происходит так называемое двойное оплодотворение, составляющее особенность цветковых растений. Честь его открытия в конце XIX столетия принадлежит нашему русскому ученому С. Г. Навашину.
Оплодотворенное вторичное ядро начинает быстро делиться. В результате зародышевый мешок заполняется массой клеток, содержащих питательные вещества (крахмал, масло). Эту используемую для питания зародыша ткань называют эндоспермом. Оплодотворенная яйцеклетка — зигота начинает расти и делиться, в результате чего формируется зародыш, представляющий собой маленькое растение, состоящее из семядолей (двух или одной), подсемя-дольного колена и корня.
Семяпочка тем временем превращается в семя, ее покровы твердеют и образуют кожуру семени (рис. 16). Стенки завязи (плодолистики) разрастаются, становятся сочными или твердыми, кожистыми или деревянистыми. Теперь завязь превращается в плод, надежно защищающий семена. Плоды разносятся животными или ветром, и после разрушения стенок (околоплодника) семена освобождаются. Семя в благоприятных условиях прорастает и дает новое бесполое поколение цветкового растения. Таким образом, в цикле развития покрытосеменных растений также преобладает бесполое поколение.
Как мы уже говорили, низшим растениям, а также мхам и папоротникообразным для полового процесса необходима вода, в которой сперматозоиды активно движутся к яйцеклеткам. Эти растения произрастают или в воде (водоросли), или во влажных, тенистых местах (мхи, папоротники, хвощи, плауны). Половой процесс у семенных и особенно у цветковых растений не связан с водой, недостаток которой так остро ощущается при жизни на суше. Мужские гаметы (спермин) у них доставляются к яйцеклеткам при помощи пыльцевой трубки. Кроме того, семя надежно защищает зародыш. Благодаря этим особенностям семенные и особенно покрытосеменные растения смогли завоевать сушу. Они в настоящее время господствуют на Земле.
2i.SU ©® 2015